��ά��˾�������Ʒζ�����ѹ��DZ���� ��������ҩ��ѧ�İе�ʶ��ͻ���̽��
2025��5�£��Ĵ���֦��������ҽԺ��֢ҽѧ�Ƶ�С�������Ŷ�����Drug Des Devel Ther����һ����IF=4.7���Ϸ��������¡�Potential of Sivelestat for Pulmonary Arterial Hypertension Treatment: Network Pharmacology-Based Target Identification and Mechanistic Exploration�������о����֣� ����ά��˾���ƾ���ͨ����ͬ�е��;�����Ʒζ�����ѹ��PAH����DZ���� ������ʵ���������100mg/kg�ļ���ʹ����ά��˾���ƿ��������� PAH �����Ʒ�Ѫ�����ܡ� ������ʵ���������ά��˾��������Ч����ȱ���յ��ķζ�����Ƥϸ����PAECs����ƽ����ϸ����PASMCs������ֳ��Ǩ�ơ�
Ŀ�ģ���ά��˾������һ��������������ϸ�����Ե���ø���Ƽ���Ŀǰ�ѱ����������Ƽ��Է����˺ͼ��Ժ��������ۺ�����������ά��˾������ȷ���Ŀ��Ϳ��������ԣ������Ʒζ�����ѹ��PAH������Ч�Բ�ȷ�������о�ּ��̽����ά��˾�������� PAH ��DZ���� ������ʹ��ȱ���յ��Ĵ���ģ�ͣ�10%������4�ܣ��ͷζ�����Ƥ/ƽ����ϸ����1%������������ά��˾���ƶ�PAH��Ӱ�졣���������ά��˾���ƣ�20-100 mg/kg������ 2 �ܣ�������ѪҺ����ѧ��RVSP����Ѫ�����ܣ�%WT�������������⣬��ά��˾���ƣ�50-200 μM��������ȱ����������ֳ��CCK-8��EdU����Ǩ�ƣ�Transwell����Ѫ�����ɡ�ͨ�� qPCR/Western ӡ�����еķ�����֤֤ʵ�˹ؼ��е㣨IGF1R��JAK1��JAK2��PDGFRB������ļ��١� �����ͨ��Ԥ�����������ȷ���� 595 ������ά��˾�������� PAH ��ص�DZ�ڻ���ֵ��ע����ǣ�ERBB2��IGF1R��JAK1��JAK2��PDGFRB �� PTPN11 ��Ϊ�ؼ�����Ŧ����������ʵ���������100mg/kg�ļ���ʹ����ά��˾���ƿ��������� PAH �����Ʒ�Ѫ�����ܡ�����ʵ���������ά��˾��������Ч����ȱ���յ��� PAECs �� PASMCs ����ֳ��Ǩ�ơ� ���ۣ���ά��˾���ƾ���ͨ����ͬ�е��;������ PAH ��DZ�������dz�����������ά��˾�������� PAH �ķ��ӻ��ƣ���ͨ�����ӶԽ��о���ʵ�鷽�������˳�����֤��
�����в�ѧ�о����ƣ�PAH �ķ�����ԼΪÿ�����˿� 15-50 ������������DZ�ڲ������ط��� PAH�������֯���������ļ��������졣PAH �ķ��������Ƕ����صģ��漰���ӵķ��ӵ������磬����Ѫ�����ܡ���֢����Ƥ�����ϰ���Ŀǰ�����Ʒ�������Ѫ�����ż�����Ƥ�������������������ø-5���Ƽ���Ȼ�������������֢״�������˶������Ʋ��Ե������� PAH ���漰�ķ���ͨ·���۸��ӣ����������Ҫ�����Щ���Ƶ�����ҩ������ո��ƻ��ߵ�Ԥ������������� �µ�֤�ݱ�����������ϸ�����Ե���ø��NE���� PAH �Ľ�չ�йأ���ͨ������ϸ������ʳɷ֡��������ϸ�����Ӻ��յ���Ƥ�����ϰ����ٽ�Ѫ�����ܡ���Ϊһ�������� NE ���Ƽ�����ά��˾�������ڼ��Է����˺�ŧ��֢�ٴ�ǰģ������ʾ������Ѫ�����˺���֢��Ӧ����Ч���ؼ����ǣ�������о�ǿ���� NE ���ƶ��ζ���ƽ����ϸ�� (PASMC) ��ֳ����Ƥϸ�������е����ã���Щ������ PAH �������Ƶĺ��ġ�ͨ������ NE����ά��˾���ƿ��ܻ��ƻ���Щͨ·���Ӷ�Ϊ���� PAH �ṩһ���µĻ��ơ����е� PAH �Ʒ�����Ѫ�����ż�����Ƥ����������Ҫ���Ѫ������ѧ�쳣���������Խ��Ѫ���������⡣��ά��˾���Ƶ�˫������--������֢��ֱ������Ѫ��ϸ����ֳ--ʹ��������Ϊ���һ���ƿհĺ�ѡҩ�����ҩ��ѧ��һ��������ϵͳ����ѧ��ҩ��ѧ���·�����ʹ�о���Ա�ܹ�̽��ҩ��е�ͼ���֮�������á�����ҩ��ѧ�� 2000 ��������룬ǿ��������ϵͳ�ĸ����Ժͼ����Ķ��������ʡ����о�ּ����������ҩ��ѧ������ά��˾�������� PAH ��DZ�ڰе��������ƣ��Ӷ������˽�������ָ��Ӽ������������塣�о�����ͼ���£�ͼ 1����
ͼ 1 ����ҩ��ѧ�о�����ͼ
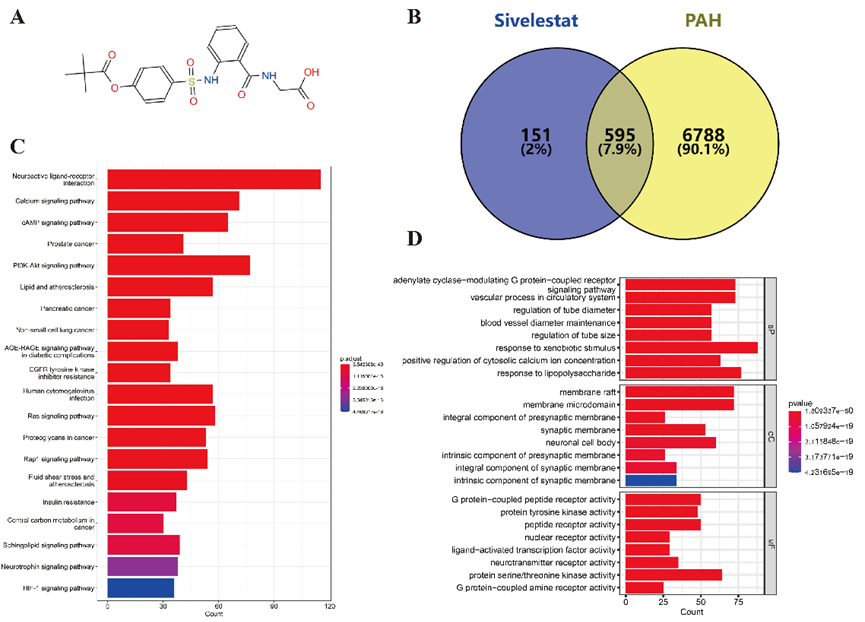
1 .��ά��˾���ƺ� PAH �б�ļ��� ��ά��˾���ƺ� PAH �б�ļ������������� ZINC ���ݿ��о�����ά��˾���ƵĶ�ά�ṹ��ͼ 2A�������������PharmMapper��SuperPred��Swiss��TargetNet���ݿ��н�����������ɾ�����ظ������ݿ⣬����ȷ����746��������Ϊ��ά��˾���Ƶ�DZ�ڰе㡣Ϊ���ռ��� PAH ��ص��²��������������� GeneCard��MalaCards �� OMIM ���ݿ⡣�����ظ������7383 ������ȷ��Ϊ��ѡ�²�����ͨ������Щ������֮ǰȷ������ά��˾���Ƶ�DZ�ڰе㽻�棬���ǵõ�����ά��˾�������� PAH �� 595 ��DZ�ڰе���ͼ 2B���������������OmicShare����ƽ̨����595�����������GO����������KEGG�ź�ͨ·�������Բ�����ά��˾��������PAH�Ŀ��ܻ��ơ�ͨ��ֱ��ͼ��ʾ�˰�qֵ������ǰ20��KEGG�ź�ͨ·�Լ�ǰ8��BP��CC��MF���ͼ��չʾ�˶�ǰʮ�� KEGG �ź�ͨ·�ķ�������Щͨ·���� ��������-��������á����ź�ͨ·��cAMP �ź�ͨ·��ǰ���ٰ���PI3K-Akt �ź�ͨ·��֬���붯������Ӳ�������ٰ�����Сϸ���ΰ���������֢�е� AGE-RAGE �ź�ͨ·�Լ���Ƥ�������������Ұ��ἤø���Ƽ���ҩ�ԣ�ͼ 2C����ǰ�˸� BP ������������ỷ��ø���� G ����ż�������ź�ͨ·��ѭ��ϵͳ�ڵ�Ѫ�ܹ��̡��ܾ����ڡ�Ѫ��ֱ��ά�֡��ܾ����ڡ�����������̼��ķ�Ӧ��ϸ��Ĥ������Ũ�ȵ���������Լ���֬���ǵķ�Ӧ����ص� CC �������Ĥ����Ĥ��ͻ��ǰĤ�������ɷ֡�ͻ��Ĥ����Ԫϸ���塢ͻ��ǰĤ�Ĺ��гɷ֡�ͻ��Ĥ�������ɷֺ�ͻ��Ĥ�Ĺ��гɷ֡���ص� MF ������� G ����ż����������ԡ������Ұ��ἤø���ԡ���������ԡ���������ԡ����弤���ת¼���ӻ��ԡ�����������ԡ�����˿����/�հ��ἤø���ԡ�G ����ż����������ԣ�ͼ 2D����
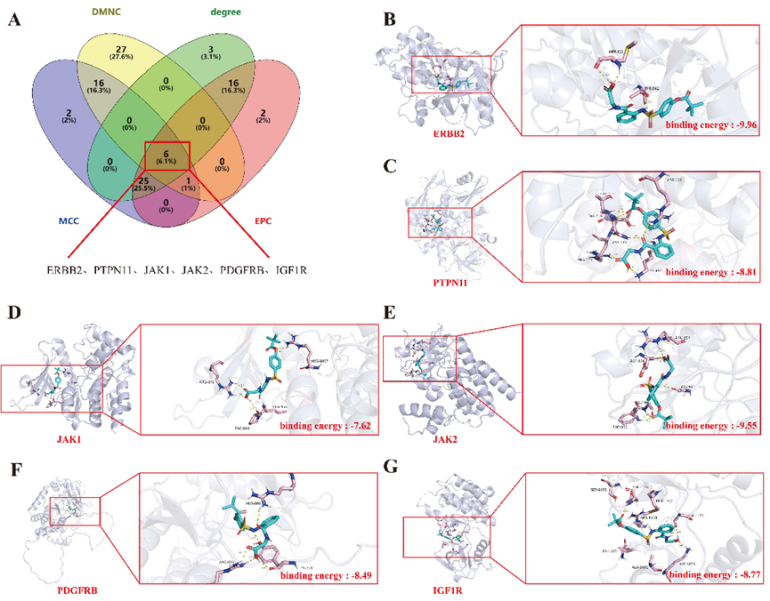
ͼ 2 ��ά��˾�������� PAH ��DZ������е㡢���ܹ��̺��ź�ͨ·��(A) ��ά��˾���ƵĻ�ѧ�ṹ��(B) ͨ��ά��ͼȷ����ά��˾�������� PAH �� 595 ��DZ�ڰе㡣(C) KEGG ���������ó���ǰ 20 ���ź�ͨ·��(D) ͨ�� GO ���������ó���ǰ 20 ��������� (BP)��ϸ���ɷ� (CC) �ͷ��ӹ��� (MF)�� 2 .ͨ�����ӶԽ��о�ȷ����ά��˾��������Ŧ����Ľ������ Ϊ��ȷ������ά��˾�������� PAH ��ص���Ŧ�������Dz����������㷨�� �ȡ�MCC��EPC �� CMNC���Ӷ�ȷ����ǰ 50 ���е㡣���У����������������ص��е㣺 ERBB2��PTPN2��JAK1��JAK2��PDGFRB �� IGF1R��ͼ 3A������������Ŧ����ܿ�������ά��˾���ƶ� PAH ������Ч���з�����Ҫ���á����Ǵ� PDB ���ݿ��м���������Щ��Ŧ�������ά����ṹ�������������Autodock Vina���������˺��Ļ�������л���֮��ķ��ӶԽӣ�������Pymol�����ԶԽӽ�������˿��ӻ��������˽���ܡ��Խӽ�����������ɽ���ܳ���-6.5 kcal/mol��������������ȶ���������˵��ERBB2����ά��˾���ƵĽ����Ϊ-9.96 kcal/mol��ͼ3B���������PTPN2Ϊ-8.81 kcal/mol��ͼ3C����JAK1Ϊ-7.62 kcal/mol��ͼ3D����JAK2Ϊ-9.55 kcal/mol��ͼ3E����PDGFRBΪ-8.49 kcal/mol��ͼ3F����IGF1RΪ-8.77 kcal/mol��ͼ3G����
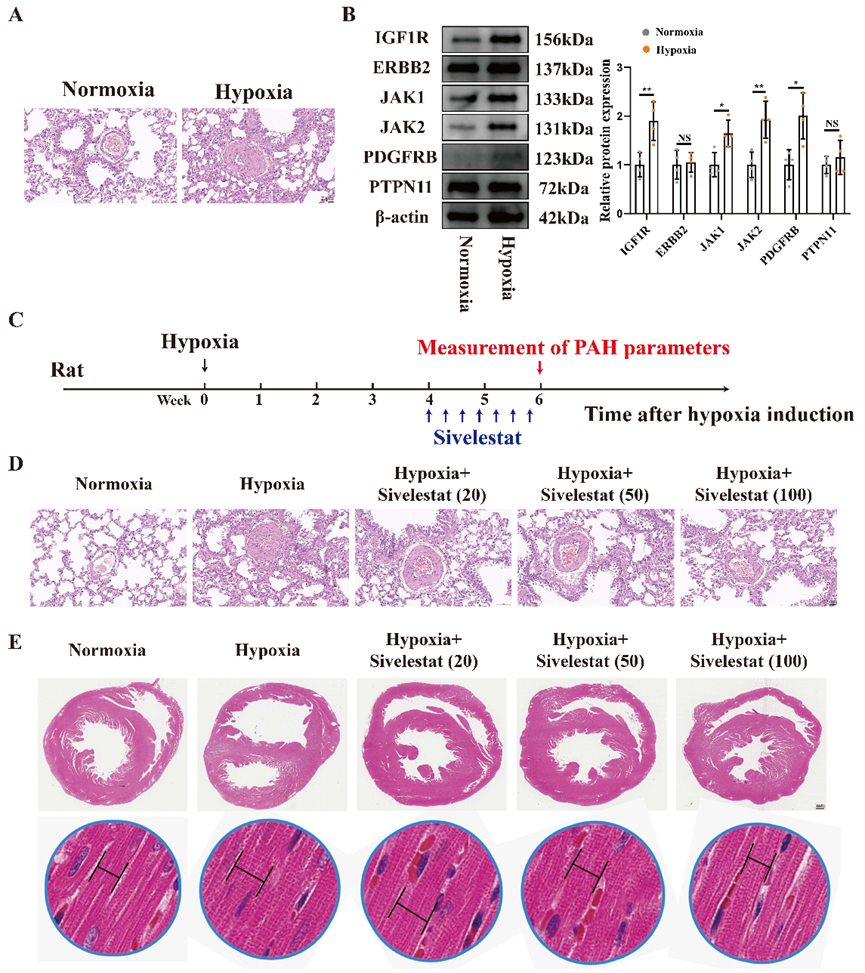
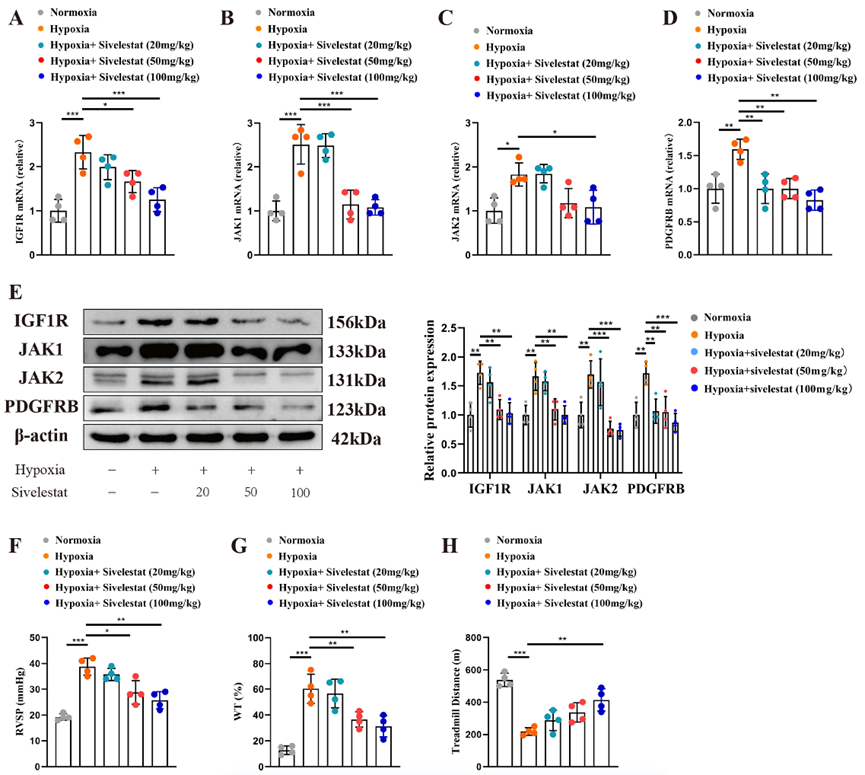
ͼ 3 ��������ҩ��ѧ����������ά��˾�������� PAH ��DZ�ڰе㡣(A���ռ��� Degree��MCC��EPC �� DMNC �㷨������ǰ 50 ���������ص��İе�ʶ��Ϊ���Ļ���(B-G�����÷��ӶԽ���֤����ά��˾��������Ŧ����Ľ�ϣ���������Ӧ�Ľ���ܡ� 3 .��ά��˾�����ܻ���ȱ���յ��� PAH ��Ѫ�����ܺ������ҷʴ� Ϊ���о���ά��˾�����Ƿ���ͨ��Ӱ��������Ŧ���������� PAH�����ǽ�����һ�������յ��� PAH ģ�ͣ�ͼ 4A����Ŀǰ���о���������ά��˾���ƾ��п��ͱ���Ѫ����Ƥ�����ԡ����������ERBB2 �� PTPN11 �� PAECs �еı���ˮƽ��Ա��ֲ��䣬�� IGF1R��JAK1��JAK2 �� PDGFRB �ı������������ӣ�ͼ 4B����������Ϊ�������������ĸ���ԭ���ǣ�ERBB2 �� PTPN11 �� PAH �Ľ�չ�����в����� PAECs �з��ӹؼ����ã��෴�������� PASMCs ������ϸ�������п��ܸ�������ԡ���ˣ���ά��˾���ƽ鵼����Ƥ�������ÿ�����Ҫ��ͨ�� IGF1R��JAK1��JAK2 �� PDGFRB �ٽ��ġ�Ϊ���������ǵļ��裬������������¶���ʵ�鷽����ͼ 4C��������������볣������ȣ�ȱ�����Ѫ��ֱ������ǻ��խ���� 100 mg/kg�ļ����£���ά��˾��������������Ѫ��ֱ����ǻ��խ�ı仯��ͼ 4D����ͬ����������ȱ������ȣ�ȱ���������ҵ��ļ�ϸ����������ͷʺ�ֵ��ע����ǣ�������ά��˾����Ũ�ȵ����ӣ������ҷʴ������Ը��ƣ��ļ�ϸ���ʴ���٣�ͼ 4E����
ͼ 4 ��ά��˾�����ܻ����Ѫ�����ܺ������ҷʴ�(A) ȱ���ͳ��������·�Ѫ�ܵĴ�����ͼ������ͼ��20 �ס�(B) ȱ���ͳ��������´��� PAECs �и��ֵ����ʵı���ˮƽ��(C) ����ʵ������ͼ��(D) ��ͬ���������·�Ѫ�ܵĴ�����ͼ������ͼ��20 μm��(E) ��ͬ���������µ�������Ƭ�����ࣺ1 ���ס�*P < 0.05, **P < 0.01, ***P < 0.001, ns�� �������ԡ� 4 .��ά��˾���ƿ�ͨ������ IGF1R��JAK1��JAK2 �� PDFRB �ı��ﻺ�� PAH Ϊ�˲�����ά��˾���ƻ����Ѫ�����ܵľ�����ƣ����Ƿ����˸������ķ�Ѫ�ܣ��γ�Ѫ����Ĥ����������IGF1R��JAK1��JAK2��PDFRB��mRNA�͵��ױ���ˮƽ������������볣��������ȣ�ȱ������������ PAECs �� IGF1R��JAK1��JAK2 �� PDFRB �� mRNA ������֮�£���ά��˾���������Խ�����Щ�����ı���ˮƽ��ͼ 5A-D�����ڵ�����ˮƽ��Ҳ�۲쵽��һ�µĽ����ͼ 5E����Ϊ��ȷ���� PAH �Ļ���̶ȣ����Dz����� RVSP��ֵ��ע����ǣ���ά��˾�����������ܵ���������������ȱ�������RVSP���ߣ�ͼ5F�������Dz��� WT% ������Ѫ�����ܣ�ͬ������ά��˾����Ҳ������Ѫ�����ܵĽ����ͼ 5G����������̬ѧ�仯�����ǻ��о��������仯�����ǶԴ����˶������IJ��Ա�����ȱ���������˶����������½���Ȼ������ά��˾�������ƺ�����˶��������������ͼ 5H��
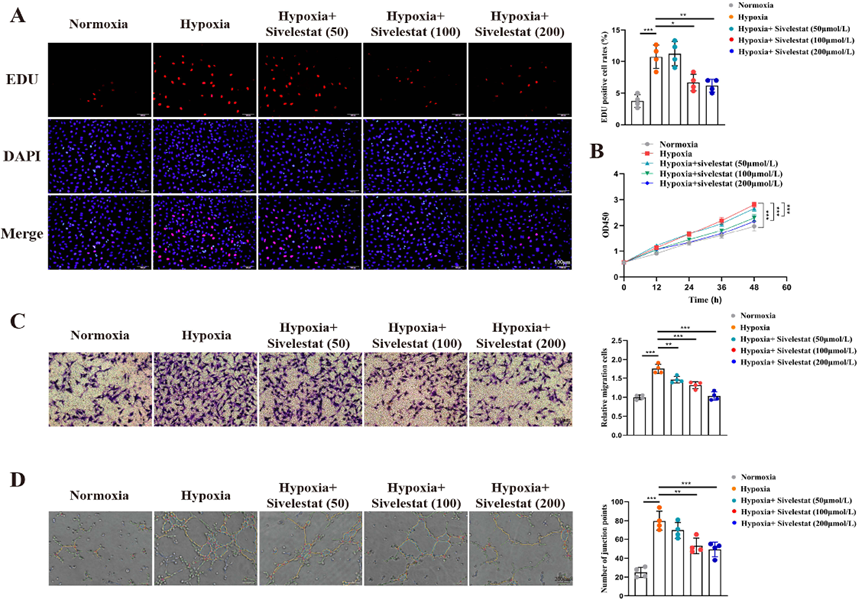
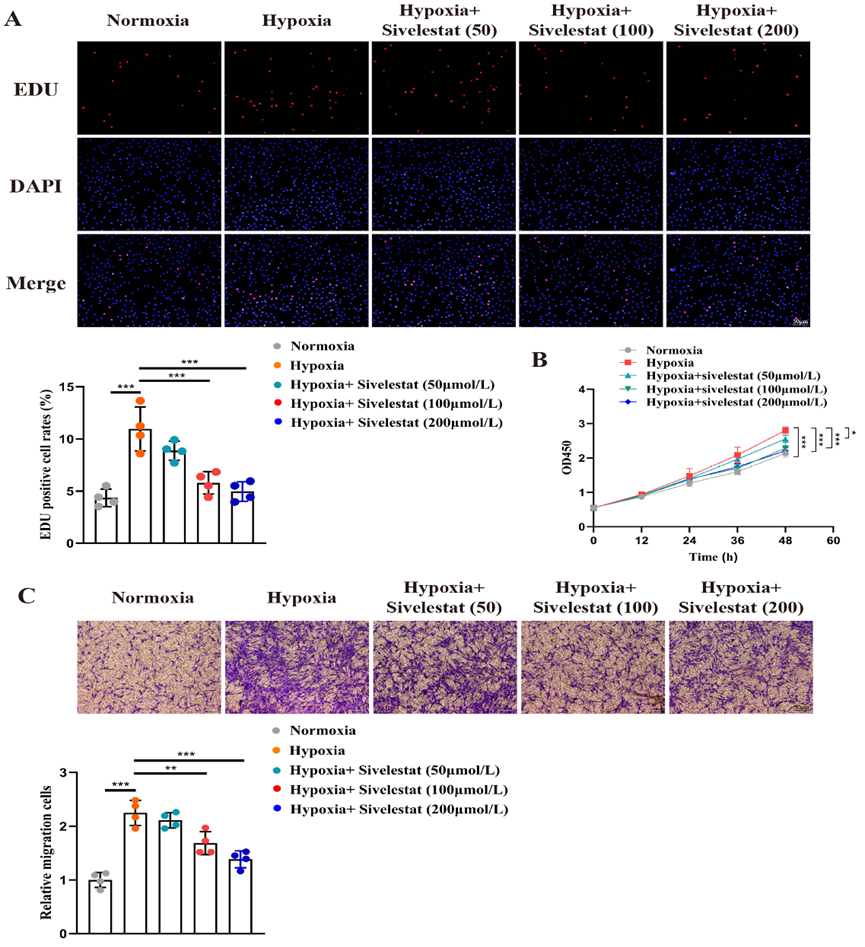
ͼ 5 ��ά��˾���ƻ��� PAH �Ĺ��̡�(A-D����ͬ���������´����Ѫ����Ƥϸ���� IGF1R��JAK1��JAK2 �� PDGFRB �� mRNA ����ˮƽ��(E����ͬ���������´����Ѫ����Ƥϸ���� IGF1R��JAK1��JAK2 �� PDGFRB �ĵ��ױ���ˮƽ��(F) ��ͬ���������´����������ѹ����(G����ͬ���������´���Ѫ�ܱں�ȱȡ�(H����ͬ���������´�����˶�������*P < 0.05��**P < 0.01��***P < 0.001�� 5 .��ά��˾���ƿɼ���ȱ���յ��� PAECs ��ֳ��Ǩ�ƺ�Ѫ������ PAECs �Ĺ�����ֳ��Ǩ�ƺ�Ѫ������������ PAH ��������Ҫԭ��Ϊ�˽�һ��������ά��˾���Ƹ���Ѫ�����ܵĻ��ƣ������о�����ά��˾�������ƺ� PAECs �ı��ͱ仯�����Dz��� EDU �� CCK8 ��ⷨ���� PAECs ����ֳ������EDU ���������ȱ�������ٽ��� PAEC ����ֳ������ά��˾�����������ȱ������ֳЧӦ��ͼ 6A����CCK8 ����Ҳ֤ʵ����һ���֣�ͼ 6B�������⣬���ǻ����� Transwell ���������� PAECs ��Ǩ�����������������ȱ����ǿ�� PAEC ��Ǩ������������ά��˾���ƴ����������ȱ���յ���Ǩ����ͼ 6C����������ά��˾���Ƶ����ðе���Ѫ���γ�������أ�ͼ 3D��������ͨ����Ѫ���γ�Ϊ�ص��ʵ��Դ˽����˽�һ��������������֣�ȱ���̼� PAEC Ѫ���γɣ�����ά��˾��������������ȱ���յ���Ѫ���γ���ͼ 6D����Ϊ�˲��� IGF1R��JAK1��JAK2 �� PDGFRB �ڽ鵼��ά��˾���������еĹؼ����ã����ǹ���������Щ�б꣨ͼ S1�������ǹ۲쵽�������� IGF1R��JAK1��JAK2 �� PDGFRB ������ת��ά��˾���ƶ�ϸ����ֳ��Ǩ�Ƶ��������á�ֵ��ע����ǣ�JAK2����������ά��˾���ƻ��Ե�������Ϊ���ԣ�ͼ S2����
ͼ 6 ��ά��˾���ƿɼ���ȱ���յ��� PAECs ��ֳ��Ǩ�ƺ�Ѫ�����ɡ�(ͼ 6 ��ά˾������ȱ���յ��� PAECs ��ֳ��Ǩ�ƺ�Ѫ�����ɡ�����ͼ��100 �ס�(B���ڲ�ͬʱ������ CCK8 ��⣬������ͬ���������µ�ϸ��������������ϸ����ֳ���ߡ�(C������ Transwell ����������ͬ���������µ�ϸ��Ǩ��������ϸ������Խ�࣬Ǩ������Խǿ������ͼ��100 �ס�(D) ���ù��γ�����������ͬ����Ѫ����������������� ImageJ ��������������ͼ��200 μm��*p < 0.05��**p < 0.01��***p < 0.001�� 6 .��ά��˾���ƿɼ���ȱ���յ��� PASMCs ��ֳ��Ǩ�� ���ǵĹ۲��������Ѫ�����ܵ���֯ѧ��̬��ʾ��PASMCs ����ֳ��Ǩ���ǵ��¹�ǻ��խ�ܱ��������Ҫ���ء���ˣ����ǽ�һ���о�����ά��˾���ƶ���Щϸ����Ӱ�졣EDU ʵ����������ȱ����������ǿ PASMCs ����ֳ��Ȼ����ʩ����ά��˾���ƻ��������ȱ����ϸ����ֳ��Ӱ����ͼ 7A����CCK8 ʵ��Ҳ�ó������ƵĽ��ۣ�ͼ 7B�������⣬���ǻ�ʹ�� Transwell ���������� PASMC ��Ǩ�����������������ȱ���ٽ�����Щϸ����Ǩ�ƣ�����ά��˾���Ƶ�ʩ������Ч���������ִٽ�������ͼ 7C�������⣬������ IGF1R��JAK1��JAK2 �� PDGFRB ����ת��ά��˾���ƶ� PASMC ��ֳ��Ǩ�Ƶ��������ã����й����� JAK2 �����Ч�������ԣ�ͼ S3����
ͼ 7 ��ά��˾���ƿɼ���ȱ���յ��ķζ���ƽ����ϸ����ֳ��Ǩ�ơ�(A���� EdU ��ⷨ������ͬ���������� PASMC ����ֳ��������ɫ��ʾ��ֳϸ������ɫ��ʾϸ���ˡ�����ͼ��50 �ס�(B���ڲ�ͬʱ������ CCK8 ��⣬������ͬ���������µ�ϸ��������������ϸ����ֳ���ߡ�(C������ Transwell ����������ͬ���������µ�ϸ��Ǩ��������ϸ������Խ�࣬Ǩ������Խǿ������ͼ��50 μm��*p < 0.05��**p < 0.01��***p < 0.001
������ƫ�RMSD�����������������帴���ﹹ���ȶ��ԵĿɿ�ָ�꣬������ԭ�����������ʼ���͵�λ��ƫ�RMSD ֵԽ�ͣ�˵�������ȶ���Խ�á���ˣ�RMSD��������������ģ��ϵͳ��ƽ��״̬��JAK1-Sivelestat �������� 60 �����ﵽƽ�⣬RMSD �����ȶ��� 3.0 Å ���ң�IGF1R-Sivelestat �������� 20 �����ڴﵽƽ�⣨RMSD ~3.4 Å������ PDGFRB-Sivelestat ϵͳ�� 80 �����ﵽƽ�⣨RMSD ~9.4 Å����ֵ��ע����ǣ�JAK2-JH1-��ά��˾���Ƹ������� 10 ������Ѹ���ȶ���RMSD ~2.9 Å����ͼ S4A������Щ�����ͬ��������ά��˾������е��ף�JAK1��IGF1R��PDGFRB �� JAK2��֮��Ľ���ȶ��Ժ�ǿ�� ��һ���Ľṹ�����������ڷ��Ӷ���ѧģ������У����и�����Ļ����뾶��Rg���Ϳ��ܽ�������SASA��ֵ������������ͼ S4B-S4C����������������뵰��������ù����д��ڹ��������ͬʱ����������ṹ�������ԡ� ����ڽ鵼����-����������ú��ȶ�����ʶ�������Źؼ����á����������Ӷ���ѧģ������У�����ϵͳ�ط�������ά��˾��������ֵ����ʰб�֮��Ķ�̬�������ͼ S4D���� JAK1 - ��ά��˾���Ƹ���������������ΧΪ 0 �� 6��������ģ��켣����Ҫռ��Լ 4 �������ͬ����IGF1R-��ά��˾����ϵͳ�������Ҳ�� 0 �� 6 ֮��仯�����ڴ����ȡ�������У����ƽ�������Ը��� 5���� PDGFRB-Sivelestat �������й۲쵽�˸����Եı仯������� 0 �� 8 ���ȣ�����Ҫ�������״̬�����˴�Լ 4 �������JAK2-JH1-Sivelestat ������������Χ���� 0-7 ֮�䣬�ڸ��г�Ч�Ľ�ϽΣ������ģֵ̬Ϊ 4�� ������������RMSF��������ͼ S4E����ʾ�����и�����IJл�����Զ������ޣ������������л��IJ��������� 3.0 Å�� ��֮��ȫ��ķ��Ӷ���ѧ����֤ʵ����ά��˾�������ĸ��Ұ��ἤø�б꣨JAK1��IGF1R��PDGFRB��JAK2��֮���ȶ��Ľ�Ϲ������ص����������������á����Ľṹ�����ͳ����Ĺ����ȶ��ԡ���Щ���㷢��������֧������ά��˾����ͨ��ǿ��ķ���ʶ����Ƴ�Ϊ��е����Ƽ���DZ����
�ڱ��о��У����Dz�������ҩ��ѧ����Ԥ�Ⲣͨ��ʵ����֤������ά��˾�������� PAH ��ص�DZ������е㡢���ܹ��̺��ź�ͨ·�����⣬���ǻ�ͨ��ʵ����֤ȷ���˰���IGF1R��JAK1��JAK2��PDGFRB���ڵĹؼ�����е㡣���ǵ��о������������ά��˾���ƿ�ͨ������ PAECs �� PASMCs ����ֳ��Ǩ���Լ����Ʒ�Ѫ������������ PAH�� ��������ҩ��ѧԤ�⣬����ȷ����������Ŧ����Ȼ����Ŀǰ���о���������ά��˾��������Ƥ������������Ҫ���á���ˣ�����ר���� PAECs �н�������֤�������ĸ��������˱仯�������Ʋ���һ������������ڲ��������� PAH �����йصĻ�������Ƥϸ���з������ã�����Ҳ����ͨ���漰ƽ����ϸ��������ϸ���ͳ���άϸ����;��Ӱ�� PAH������û�н�һ��̽���ⷽ������⣬��Ҳ�������о��ľ�����֮һ�� ���⣬����ͨ������ PI3K/AKT/mTOR �ź�ͨ·�ڼ����ķ��������з������á����ǵ�ʵ��֤ʵ PAH ���� PAECs �� IGF1R �����ϵ���ֵ��ע����ǣ����Ƿ���ʹ����ά��˾���ƿ����� IGF1R �ı���������ά��˾����ͨ������ IGF1R ���������� PAH�� JAK1 �� JAK2 ��ϸ�����Ұ��ἤø���� JAK-STAT �ź�ͨ·�з��ӹؼ����ã��� JAK-STAT �ź�ͨ·���ڽ鵼����ϸ����Ӧ��������֢�����߹���������Ҫ������ JAKSTAT �ź�ͨ·�������� PAH����Ϊ���ܸı�ò��IJ������̡��о����֣����� JAK1 �� JAK2 �ܷ�ֹ��Ѫ�ܴ���ά�������� PAH�����⣬��ά��˾�������������� JAK1 �� JAK2 �ı�������������ͨ������ JAK-STAT �źŴ������ڻ��� PAH ���̽�չ���淢���������á� PDGFRB ��һ�������Ұ��ἤø���ڵ���ϸ����ֳ������Ѫ�����ܷ��淢������Ҫ���á��������ά��˾���ƿ���ͨ���� PDGFRB �ź�ͨ·������öԷζ�����ѹ��������Ӱ�죬���������ڼ���Ѫ�����ܺ���Ԥ�� �� IGF1R��JAK1/JAK2 �� PDGFRB ȷ��Ϊ��Ŧ����ͼ 2-3���������� PAH ���������еļȶ����ã��� IGF1R/PASMC ��ֳ��42 JAK-STAT �ź���Ѫ�������е�����49����һ�¡���Щ�е�ķ��ӶԽӺ�ʵ�����ƣ�ͼ 5-7����֤����������ά��˾���ƻ��Ƶ�����ԡ��۲쵽�� RVSP��Ѫ�����ܺ� PAEC/PASMC ��ֳ/Ǩ�Ƶļ��٣�ͼ 4-7��ֱ��֧������ά��˾����ͨ������ȱ������;������ PAH �Ľ��ۡ� ���о�������ҩ��ѧ�����ʵ�����ϣ�����̽������ά��˾�������Ʒζ�����ѹ�Ŀ����ԣ����ֳ��������ơ������״η�������ά��˾������Ϊ�ζ�����ѹ��DZ������ҩ�����ʾ������ܵ����ڻ��ơ���Ȼ���о���������ά��˾����ͨ��IGF1R��JAK1/JAK2��PDGFRB���ڻ���ȱ���յ���PAH��DZ�������м���������ֵ�ÿ��ǡ�ȱ��ģ����Ȼ���㷺ʹ�ã�����������ȫ������ PAH ��ص�������ϸ�����Ե���ø��������֢ͨ·������ҩ��ѧȷ���˿���ҩ�е������˳���ų��˵��͵� PAH ������ BMPR2��������������С������ͳ����������Ȼ����ҩ��ѧΪȷ����ά��˾������ PAH �е�DZ�ڰе��ṩ��һ���㷺�Ŀ�ܣ������ַ���������ǿ�����ǹ�ȶ�����ȡ����о����Ƚ�ʾ�˶�е�����ã��� IGF1R��JAK1/JAK2��PDGFRB�����Է�ӳ PAH �Ķ��������ʣ�����δ��ȫ���������������أ���ת¼���ӡ���ø������Щ�е�֮��ķּ��źŹ�ϵ��δ�����о�Ӧ���϶���ѧ���ݣ���ת¼��ѧ�����ữ������ѧ�����Խ���������β���֤��ά��˾���Ʒ�������Ч������Ҫ;����
1. �����뷽�� ��ļ2021��12����2022��6���ڼ��ں���ʡ�ؿ�ҽԺ�����Ҽ������������Ƶ�AAAD ����72��������������ֱ��������Ϊ��ά��˾������(S��)����˾������(U��) ��ÿ��36��������������ϱȽϲ�����ͳ��ѧ����(P��0.05)������1����һ��δ���뻼���ٴ��������о��߶��о����顢��Ԥ�ͽ�����м�¼�����ߡ�����Χ���ڹ���������ҽʦ�����ҽʦ����������Ա���Է��顢��Ԥ������֪�顣ָ��۲��ߺ�ͳ��ѧ�����߶Է��顢��Ԥ������֪�顣���о������ʡ�ؿ�ҽԺҽѧ����ίԱ����������: (2021)�����(10-04)�ݣ��о����������ǩ��֪��ͬ���顣 2. ����������-����������ã�PPI������ �ڱ��о��У����ǽ���������� String ���ݿ⣬��ʹ�����˵��ض����ý��з��������ǽ����Ŷ���ֵ�趨Ϊ���� 0.9����ѡ�����ضϿ��Ľڵ㣬�Ӷ����ɻ��� PPI ���繩��һ�������� 3. ��Ŧ����ľ������ �ڶ� PPI ���ݽ�������������������� Cytoscape �����е� MCODE �� Cytohubba ����ֱ����ɽڵ�����ʶ�����Ľڵ�27,28��MCODE ���ר������ʶ�������������ѧ���ܵĽڵ�أ�ÿ�����е÷���ߵĽڵ��Ϊ SEED������Ϊ���ض��صĹؼ�Ŀ�ꡣ���⣬Cytohubba ������ṩ 11 �ֲ�ͬ�����˷������������� PPI �����нڵ�����������������29 ��Ŀ����ʶ�����е���Ŧ����Ϊ�����ʶ�����Ŀ��Ŷȣ����Dz����˶��������������ر��������㷨�� Ϊ�����ʶ�����Ŀ��Ŷȣ����Dz����˶��������������ر��������㷨���ȣ�ֱ������õ��������������������ԣ�MCC�����ȫ�����������еĽڵ㣩����Ե���ɷ֣�EPC����Եȥ���ĵ��ԣ�������ڽ��ɷ��ܶȣ�DMNC���ֲ������ܶȣ�����ȷ����Ҫ�Ļ���29,30�����յ����Ļ��� ERBB2��IGF1R��JAK1��JAK2��PDGFRB��PTPN11 �����������㷨�о�����ǰ 50 λ����˱����ȿ��ǡ� 4. GO �� KEGG ͨ·�������� ���ܸ����������� OmicShare ��һ����ѵ��������ݷ���ƽ̨��רΪ���� GO �� KEGG ͨ·������������ơ�31 �� GO �����У��۲쵽������������̣�BP����ϸ����֣�CC���ͷ��ӹ��ܣ�MF���������������������ͬʱ��KEGG ͨ·����ȷ�������о�������صĹؼ��ź�ͨ·���÷��������� q ֵ����������ˮƽ���ٽ���ֵ�趨Ϊ p < 0.05�� 5. ���ӶԽ� ���ȣ����Ǵӵ��������ݿ⣨PDB���л�ȡ�������ؼ��������ά����ṹ��33 ������Ǵ� PubChem ���ݿ��л�ȡ����ά��˾���Ƶ���ά�ṹ��34 ��ʹ�� OPEN Babel ���������ʽת��Ϊ PDB �ļ�������ֵС�� 6.5 kcal/mol ��ʾ��������б�֮��Ľ����Ч�� 6. ���Ӷ���ѧģ�� ʹ�� Gromacs 2022 �����Ը���������� 100ns ���Ӷ���ѧģ�⣨MD����ѡ�� Charmm 3636 ��Ϊ������������ѡ�� Gaff2 ��Ϊ����������ѡ�� TIP3P ˮģ��Ϊ������-����ϵͳ�����ܼ���������һ�����ڱ߽�Ϊ 1.2 nm ��ˮ��37��Ȼ��ģ���� 100,000 �����µ��������ƽ��͵��µ�ѹ����ƽ�⣬��ϳ���Ϊ 0.1 ps������ʱ��Ϊ 100 ps�����»��Ϳ�������þ�ʹ�� 1.0 nm �ض�ֵ���м��㡣����ں��£�310 K���ͺ�ѹ��1 bar�������£�ʹ�� Gromacs 2022 ��ϵͳ������ģ�⣬��ģ���� 100 ns�� 7. ����ʵ�� �ڶԴ���ʵʩ������֮ǰ���������ȸ�����ǻע�� 40 mg/kg����ͱ����ƽ�������������Ҳྱ�⾲����Ԥװ����ͭ�� PE-50 ���ܣ��й��ɶ�̩�˹�˾�����������ҡ����ܵ���һ�����ӵ�ѹ�����������й��ɶ�̩�ˣ��������о������¼���������Բ�������������ѹ��RVSP����λ��������������¼�� RVSP ֵ�ɼ�ӷ�ӳ�ζ�����ѹ���ı仯��ȡ���ķ���֯�����ǽ���Щ�걾�����ۼ�ȩ�У���������ľ ��������Ⱦɫ�������������Һͷζ��������������Ȼ�������ù�ʽ�����в�ں�ȵİٷֱȣ�WT%���� WT% = 2 ×��WT/�⾶��×100�� 8. ��ά��˾���Ƶļ����� ����ͨ������ʵ����������ά��˾���ƶ� PAH ������Ч����ͨ�����ܵ�ȱ����¶��������ζ�����ѹ��PAH��ģ�ͺ����ٽ�����ά˾���Ƹ�ҩ���ܡ�ȱ������ÿ�츹ǻע����ά��˾���ơ���ά��˾���Ƶļ�����20 mg/kg/�졢50 mg/kg/��� 100 mg/kg/�죩�Ǹ�����ǰ���о����ȷ���ġ� 9. ȱ���յ� PAH ����ģ�͵Ľ��� �ڱ��о��У�����ʹ����6��8�ܴ������Sprague-Dawley��SD���������д�������Խ���ҽ�ƴ�ѧ������״�����á�����������������ϱ���ʳ���ˮ�Ļ�ȡ�������ơ����ж���ʵ��ͳ����������ҽ�ƴ�ѧ�о�����ίԱ���������ţ�2022031001�������� 7 �����Ӧ�ں��о�С�齫ȱ��ģ�ʹ��������䱸�������������ϵͳ��Oxycycler Model A84XOV��BioSpherix, Ltd., China���Ļ����У�����Ũ�ȱ����� 10% ± 0.5%��������������Դ���������������������21% O2����������Ϊȷ��ʵ�����ڵĶ�����̼Ũ�ȱ����� 3% ���£�ʵ�����ڷ������Ƹ������ն�����̼��ͬʱʹ�ñ�ɫ�轺������ˮ������ȱ����¶4�ܺ�����ͨ����ǻע����ά��˾���ƻ�������ˮ�Դ���������ơ�������ʵ������У����д����������¶�Ϊ 23°C �� 26°C �Ļ����У��ⰵ����Ϊ 12 Сʱ��ÿ�ܶԶ���Ľ���״������Ϊ���м�⣬��ȷ��ʵ���˳�����У��о�����ǰû�д��������� 10. �˶�������� Ϊ������������˶��������������������ܲ����ϵ�����ܲ����롣���Կ�ʼʱ���ܲ������ij�ʼ�ٶ��趨Ϊ 10 ��/���ӣ�ÿ 5 �������� 5 ��/���ӣ�ֱ���ﵽ 25 ��/���ӵ�����ٶȡ�Ȼ����һ����ٶȱ��� 15 ���ӡ�����Ҫô�� 30 ���Ӻ������Ҫô�ڴ�����ֳ�ƣ�ͼ���ʱ������ƣ�ͼ���Ķ����Ǵ����ڳ�������ϲ������� 5 ���ӡ����о����϶���ʵ���������� 11. ϸ������ PAECs ��Ŀ¼�� 3100���� PASMCs��Ŀ¼�� 3110������ Sciencell ��˾����ר����������Ŀ¼�� 1001��1101��������������ʵ��ѡ���˵� 3 ���� 6 ���ϸ����Ϊ�˽�һ���о���ά��˾�������� PAH ��DZ�ڻ��ƣ�������ϸ����������ȱ��������1% O2�������� 24 Сʱ��ͬʱ��������������ά��˾���ƴ�����ϸ��Ǩ�ƺ���ֳ��Ӱ�졣��ά��˾���Ƶ�Ũ�ȷֱ�Ϊ 50 μmol/L��100 μmol/L �� 200 μmol/L����ЩŨ���Ǹ�����ǰ���о����ѡ���ġ� 12. ���� PCR (qPCR) ���������̵�˵��ʹ�� Trizol �Լ���Invitrogen ��˾���ӷζ�����Ƥ��֯����ȡ�� RNA����ȡ��ʹ�÷�ת¼�Լ��У�Transgene���� 2 μg �� RNA ��ת¼Ϊ cDNA�����ʹ�� SYBR Green PCR ���Һ���й�������������ʵʱ PCR ������ABI 7500������ qPCR ������CT ֵͨ�� Applied Biosystems 7500 ���м��ϵͳ������ã�β-����������Ϊ�ڲ���ȷ�������ȷ�ԡ������������ Primerbank ��ƣ�Sangon Biotech���й��Ϻ����ϳɡ� 13. Western ӡ�� �Ӵ������֯��С�İ���ζ������������ж϶�������ȡ��Ƥ��֯���� RIPA �ѽ��Һ�͵���ø���Ƽ��Ļ�����ѽ���Ƥ��֯������˫����ᶨ������ 10% SDS-PAGE �����Ϸ��뺬�� 30 µg �����ʵ���Ʒ��Ȼ����ת�Ƶ���ƫ������ϩ��PVDF��Ĥ�ϡ��� 5%ţѪ�������Ϻ���һ���� 4°C ���� PVDF Ĥ 12 Сʱ��Ȼ������Ӧ���ֵĶ������� 2 Сʱ��������ǿ��ѧ���ⷢ��Һ��ʹ�û�ѧ����ϵͳ�Ե������������г���ʹ�� ImageJ �����ݽ��ж���������β-����������Ϊ���ض��ա���Ӧ��һ������ �ȵ������������� 1 ���壨IGF1R��ab182408��1:1000��Abcam����Erb-b2 �����Ұ��ἤø 2��ERBB2��ab134182��1:1000��Abcam����Janus ��ø 1��JAK1��ab133666��1:5000��Abcam����Janus ��ø 2��JAK2��ab108596��1�� ѪС������������������ beta��PDGFRB��ab313777��1:1000��Abcam���������Ұ�������ø�������� 11��PTPN11��ab300579��1:1000��Abcam������������ beta��β-�������ף�ab7817��1:1000��Abcam���� 14. ϸ����ֳ���� ����ʹ�� CCK-8 ����Լ��У�K1076l��APExbio��������������ϸ��������������˵������ͬ��� PAECs �� PASMCs ���ֵ� 96 �װ��С����� 6 Сʱ��ƽ��������������������ȱ�������������� 24 Сʱ��24 Сʱ�����ڽ�����������ÿ�����м��� 10 µL CCK-8 ��Һ��Ȼ���� 37°C ������ 1.5 Сʱ�����ʹ�ò���Ϊ 450 nm ���װ��Ķ���������������ȡ����⣬���ǻ�ʹ���� 5-��Ȳ��-2′-�������գ�Edu��ϸ����ֳ����Լ��У�K1076l��APExbio�������� PAECs ��ƽ����ϸ������ֳ������������������̵�˵�����ڱ��о��У���Щϸ���ڳ�����ȱ�����������������װ��С�ÿ���� 5 × 10^4 ��ϸ����ÿ����� 1 mL Edu ���� 37°C ���� 3 Сʱ��Ϊ�˹̶�ϸ�������Ǽ��� 4% ��ۼ�ȩ 20 ���ӣ�Ȼ���� 0.5 mL �����Ӧ��Һ����ÿ���� 30 ���ӡ�Ϊȷ��ʵ�����Ŀɿ��ԣ�����ʵ�������ظ��ĴΡ���������� DAPI ��ϸ����Ⱦɫ 10 ���ӣ��Է���۲�ͺ��������� 15. ϸ��Ǩ��ʵ�� ϸ��Ǩ�Ʋ��� Transwell �ͻ��۷������������ڻ��������У�����ͬ���ϸ����ÿ�� 5 × 10^5 �����ܶȽ��ֵ� 6 ���������ϣ��������ﵽ��ϡ�Ȼ���øɾ��� 200 μL ��Һ����ͷ�ڿױ��滮��һ�����Ի��ۡ�Ȼ���� PBS ��ϴϸ�����Σ�������Ѫ�������������ʵ����������� 24 Сʱ��ʹ�õ��ù�ѧ������IX71���ձ����ְ�˹��˾������Ǩ�ƿ��ȣ����� 0 Сʱ�� 24 Сʱʱ�ɼ�ͼ���� Transwell ���飬���������̵�˵��ʹ�� 24 �װ塣�ڴ������У��� 200 μL ���� FBS ��ϸ������Һ�������� 10,000 ��ϸ����������ǻ��ͬʱ����ǻ���� 600 μL �� FBS ��������������ϸ������������ 24 Сʱ��ȡ����ǻ���� 4% ��ۼ�ȩ�̶�ϸ����Ȼ���� 1%�ᾧ����Һ��ϸ��Ⱦɫ 15 ���ӣ�����ǩ�����ȥĤ�ϱ���ķ�Ǩ��ϸ��������ù�ѧ��������Ǩ��ϸ������¼ʵ������ 16. ͳ�Ʒ��� ʹ�ù��ʷ�������G*Power 3.1��α = 0.05������ = 0.8��ЧӦ��С����������ʵ�飩����ȷ����������С������������о�ÿ�� n = 8 ֻ����ȱ��ģ������ 4 ֻ������������ʵ�� n = 4 �����������ظ���������ƽ��ֵ ± ���� (SD) ��ʾ����̬���� Shapiro-Wilk ����������������������� Levene �������ȷ�ϡ������ıȽϣ�����̬�ֲ����ݲ��÷����˫βѧ�� t ���飻������÷Dz��� Mann-Whitney U ���顣�����������̬�ֲ����ݲ��õ����ط�������� Tukey ��У�����ԷDz������ݼ����� Kruskal-Wallis �� Dunn ���顣���з�����ʹ�� GraphPad Prism 9.0������ GraphPad ������˾�����У�ͳ���������趨Ϊ P <0.05��ʵ�鷽�������ظ��ĴΣ���ȷ�����ظ��ԡ� �ο����ף� Deng X, Qiu P, Li X, Hu Y, Que Q, Zhang K, Deng T, Liu Y. Potential of Sivelestat for Pulmonary Arterial Hypertension Treatment: Network Pharmacology-Based Target Identification and Mechanistic Exploration. Drug Des Devel Ther. 2025 May 20;19:4123-4138. doi: 10.2147/DDDT.S507240. PMID: 40416794; PMCID: PMC12103203.
|
������
��վ�ؼ��ʣ� �人����֮��